Malacolimax tenellus (Müller, 1774)
Limax tenellus O. F. Müller, 1774
Typ: Pravdepodobne nezvestný.
Locus typicus: Frederiksdal, Dánsko.
Synonymika: Limax tenellus Nilsson, 1822; Limax serotinus Schrenk, 1848; Limax cereus Held, 1849; Limax collinus Normand, 1952; Limax silvaticus Dumont et Mortillet, 1852; Limax fulvus Normand, 1852; Limax cinctus Heynemann, 1862; Limax xanthius Bourguignat, 1866; ?Limax Kostálii Babor, 1900 (bibliografické dáta pozri Hesse 1926).
Opis
Rozmery: do 50 mm (konzervovaný do 35 mm).
Farba tela bledo až hnedasto citrónovožltá, niekedy s hnedastým, oranžovým alebo zelenkastým odtieňom, zriedkavejšie s naznačenými tmavšími bočnými pásikmi, ktoré môžu byť naznačené aj na štíte, tmavá hlava, tmavohnedé, tmavofialové až čierne tykadlá, pokožkové papily malé a nevýrazné, kýl nevýrazný, zaoblený, krátky. Chodidlo a okolie pneumostómu belavé. Sliz vodnatý, priesvitný, žltý. Juvenily sú svetlejšie ako dospelce.
 |
| Foto: M. Horsák |
Anatómia: Penis lopatkovitý, hruškovitý alebo krátko valcovitý, v distálnej časti s nepravidelnými rozšíreniami. Vas deferens, sa otvára laterálne na proximálnom konci penisu a vytvára polmesiacovitý, často zvlnený, výbežok. Pôsobí to tak, akoby rozšírený vas deferens prirastal k zadnej stene penisu. Táto časť môže byť tmavo pigmentovaná. Penis je malý, kratší ako polovica plášťa a o niečo dlhší ako hltan. Retraktor penisu je laterálne pripojený v proximálnej polovici penisu. Vnútri penisu je komplikovaná sústava záhybov a vačkov. Spermatéka je podlhovasto oválna, uložená na tenkom, približne rovnako dlhom spermatickom kanáliku a je spojená s penisom. Vajcovod je hrubý, rúrkovitý, kratší ako penis (Wiktor 1996).
Rožšírenie
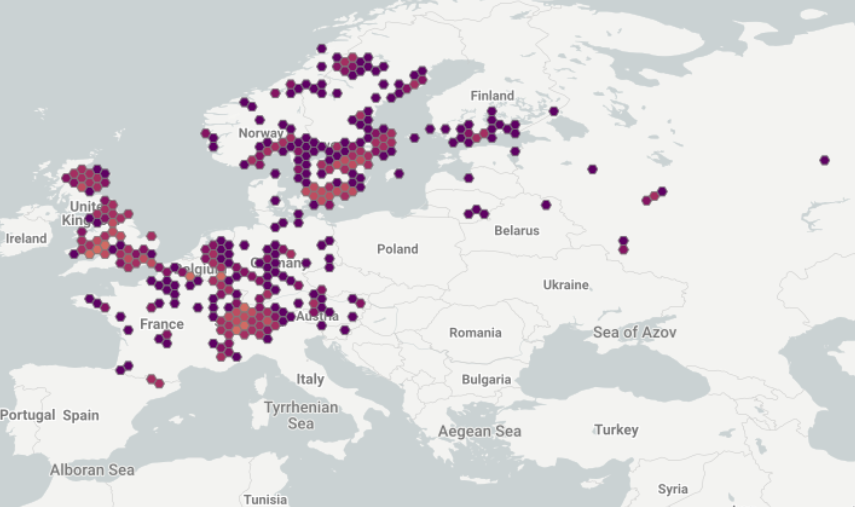
Európsky druh, s ťažiskom výskytu v západnej a severnej Európe. Druh má v Európe pomerne veľký areál, ktorý sa rozprestiera od Veľkej Británie a Francúzska na východ do európskej časti Ruska, vyhýba sa však najsevernejším a najjužnejším oblastiam. Na východe Európy sa nachádza v severozápadnej, západnej a strednej oblasti európskej časti Ruska (Sysoev a Schileyko 2009, Kantor et al. 2010) a západnej, severozápadnej a severnej Ukrajiny (Sverlova et al. 2006).
Pôvodný je v týchto krajinách: Rakúsko; Bielorusko; Belgicko; Česká republika; Dánsko; Estónsko; Fínsko; Francúzsko; Nemecko; Maďarsko; Lotyšsko; Lichtenštajnsko; Litva; Luxembursko; Holandsko; Nórsko; Poľsko; Rumunsko; Ruská federácia (európska časť Ruska); Slovensko; Slovinsko; Švédsko; Švajčiarsko; Ukrajina; Veľká Británia.
Rozšírenie na Slovensku: Ostrovčekovito, ale pomerne súvislo, je rozšírený v karpatskom bioregióne.
Ekológia a biológia
Lesný stenotopný a prevažne vlhkomilný druh. Vo Veľkej Británii sa považuje sa za indikátora starých lesov (ancient woodlands) (Kerney 1999). V niektorých častiach areálu žije aj v ihličnatých lesoch, ale je zvyčajne je menej početných populáciách ako v listnatých lesoch (Welter-Schultes 2012), kde žije najmä v rastlinnom opade pod kusmi tlejúcich driev. Toleruje tradičné extenzívne lesné hospodárenie, miestam s intenzívnym hospodárením sa vyhýba (najmä miestam s rozsiahlymi holorubmi). Vo Švajčiarsku sa vyskytuje do nadmorskej výšky 2100 m, zvyčajne však pod 1700 m (Welter-Schultes 2012).
Rozmnožovanie
K rozmnožovaniu dochádza od neskorej jesene do začiatku zimy, mláďatá sa liahnu zvyčajne skoro na jar. Obdobie rozmnožovania v októbri a novembri vo Francúzsku a Nemecku, 15 - 35 vajíčok (priemer 2 - 4 mm) sa kladie v dávkach (každých niekoľko týždňov, 4 - 6-krát) na chránených a mierne teplých miestach, mláďatá sa liahnu po 20 - 120 dňoch, mláďatá sú bezfarebné alebo svetložltkasté, žijú v podzemí a živia sa najmä hubovým mycélom, plnú žltú farbu majú po 6 mesiacoch, dospelosť dosahujú po 7 - 9 mesiacoch, potom sa v nich hromadí tmavý pigment, dĺžka života 12 - 13 mesiacov. Kopulácia je obvyklá, ale je možné aj samooplodnenie.
Potrava
Živí sa hubami, lišajníkmi a riasami, za vlhkého počasia vylieza za potravou aj na kmene stromov s relatívne hladkou kôrou (buky, hraby a pod.) (Nordsieck, weichtiere.at).
Literatúra
Hesse P (1926) Die Nacktschnecken der palaearktischen Region. Abhandlungen des Archiv fur Molluskenkunde, 2/1: 1--152.
Irikov A & Eröss ZP (2008) An updated and annotated checklist of Bulgarian terrestrial gastropods (Mollusca: Gastropoda). Folia Malacologica 16: 199–207.
IUCN (2017) The IUCN Red List of Threatened Species. Version 2017-3. Available at: www.iucnredlist.org. (Accessed: 5 December 2017).
Kantor, Y., Vinarski, M., Schileyko, A. and Sysoev, A. 2010. Catalogue of the continental mollusks of Russia and adjacent territories. Electronic version 2.3.1. Available at: http://www.ruthenica.com/documents/Continental_Russian_molluscs_ver2-3-1.pdf.
Kappes, H. 2006. Relations between forest management and slug assemblages (Gastropoda) of deciduous regrowth forests. Forest Ecology and Management 237: 450–457.
Kappes H (2017) Malacolimax tenellus. The IUCN Red List of Threatened Species 2017:
Kerney, M.P. 1999. Atlas of the land and freshwater molluscs of Britain and Ireland. Harley Books, Great Horkesley, Essex.
Seddon, M.B., Killeen, I.J. and Fowles, A.P. 2014. A Review of the Non-Marine Mollusca of Great Britain: Species Status No. 17. NRW Evidence Report. Natural Resources Wales, Bangor.
Sverlova, N.V. 2006. O rasprostranenii nekotorykh vidov nazemnykh mollyuskov na territorii Ukrainy [On the distribution of some species of land molluscs on the territory of Ukraine]. Ruthenica 16(1-2): 119-139.
Sysoev, A. and Schileyko, A. 2009. Land Snails and Slugs of Russia and Adjacent Countries. Pensoft, Sofia - Moscow.
Welter-Schultes, F. 2012. European non-marine molluscs, a guide for species identification. Planet Poster Editions, Göttingen.
Wiktor A (1996) The slugs of the former Yugoslavia (Gastropoda terrestria nuda – Arionidae, Milacidae, Limacidae, Agriolimacidae). Annales Zoologici, 46: 1–110).






.")




